
微藻光合产氢因其绿色、可再生的特性,被视为解决未来能源危机的潜在途径之一。然而,该过程的核心矛盾在于:氢化酶对氧气极度敏感,而光系统II(PSII)在分解水产生电子的同时会释放氧气。传统的缺硫诱导产氢策略虽能通过下调PSII活性来营造厌氧环境,却不可避免地限制了氢化酶所需的电子供应,导致产氢效率低下。如何在维持厌氧条件的同时,保障充足的电子流供给,是制约微藻生物产氢应用的关键瓶颈。
近日,西北大学生命科学学院付爱根/王菲团队在国际权威期刊《The Plant Journal》在线发表了一篇题为“TEF8 regulates H2 photoproduction by destabilizing PSII supercomplexes in Chlamydomonas”的研究论文。该研究鉴定到一个衣藻光合产氢的全新负调控因子—类囊体膜磷酸酶TEF8,揭示了其在缺硫诱导条件下通过调控PSII亚基的去磷酸化,进而影响PSII超复合体稳定性及光合电子流,最终实现对光合产氢效率精细调控的分子机制。

研究团队通过对多个光合作用突变体系统筛选,发现类囊体磷酸酶TEF8的缺失能够显著提升莱茵衣藻在缺硫条件下的光合产氢速率。进一步研究表明,tef8突变体在缺硫条件下能够维持更高的PSII光化学活性和电子传递效率,其PSII超复合体的稳定性显著优于野生型,核心亚基的降解受到明显抑制。这种对PSII的保护作用确保了持续的光合电子流供给,为氢化酶提供了充足的还原力。分子实验证据揭示,TEF8通过其磷酸酶活性调控PSII核心亚基D2及捕光蛋白CP29的去磷酸化,从而促进PSII在胁迫条件下的降解。当TEF8基因敲除后,PSII亚基维持高磷酸化状态,超复合体更趋稳定,电子传递得以持续。TEF8磷酸酶失活的点突变株完全重现了tef8的高产氢表型,证实了TEF8的催化活性在这一调控环节中起到核心作用(见工作模型图)。
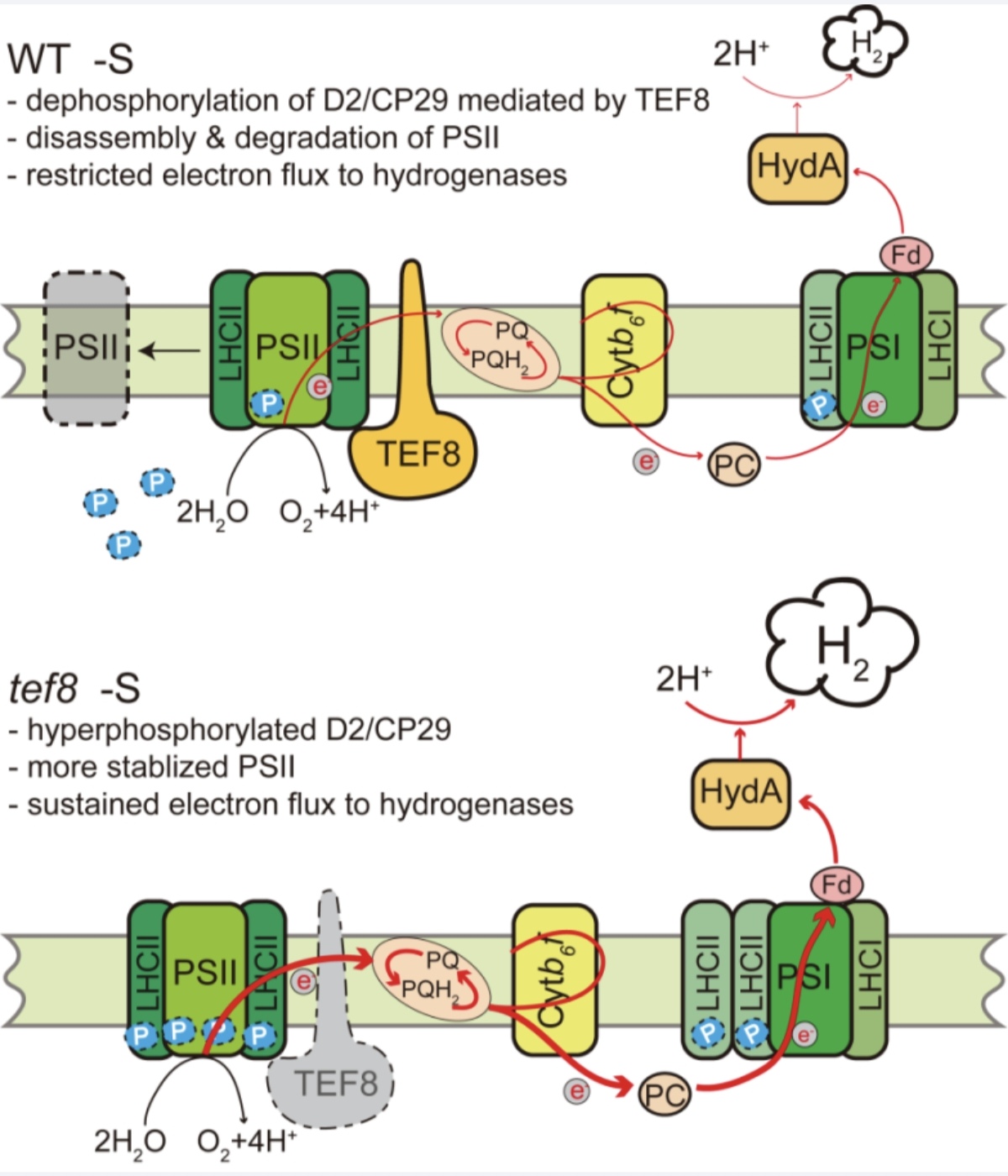
TEF8调控缺硫条件下莱茵衣藻光合产氢的工作模型
综上,该研究发现TEF8可作为PSII稳定性与产氢效率之间的分子开关,揭示了一种光合产氢调控新策略:通过抑制PSII在胁迫条件下的过度降解来维持更持久的电子输出。此工作是该团队在衣藻光合作用调控领域系列研究的延伸与深化[1-4],为未来借助合成生物学手段改造微藻、实现高效可持续的光合产氢提供了关键的理论支撑。
西北大学生命科学学院硕士研究生王小玉、博士研究生姚强为论文的共同第一作者。西北大学生命科学学院王菲副教授和付爱根教授为论文的共同通讯作者。该研究得到了国家自然科学基金委、陕西省基础科学(化学、生物学)研究院及陕西省自然科学基础研究计划的资金支持。
参考文献:
[1] Yao Q, Dong J, Zhang T, Cao H, Yu J, Wang X, Li B, Zhu L, Wang Y, Fu A, Wang F. (2025) Dimerization of immunophilin CYN38 regulates photosystem II repair in Chlamydomonas. Plant, Cell&Environment. 48: 5637-5649
[2] Dong J, Hou J, Yao Q, Wang B, Wang J, Shen X, Lai K, Ge H, Wang Y, Xu M, Fu A, Wang F. (2024) The thylakoid phosphatase TEF8 is involved in state transition and high light stress resistance in Chlamydomonas. Plant Journal. 120: 2138-2150
[3] Guo J, Yao Q, Dong J, Hou J, Jia P, Chen X, Li G, Zhao Q, Wang J, Liu F, Wang Z, Shan Y, Zhang T, Fu A, Wang F. (2024) Immunophilin FKB20-2 participates in oligomerization of Photosystem I in Chlamydomonas. Plant Physiology. 194: 1631-1645
[4] Fu W, Cui Z, Guo J, Cui X, Han G, Zhu Y, Hu J, Gao X, Li Y, Xu M, Fu A, Wang F. (2023) Immunophilin CYN28 is required for accumulation of photosystem II and thylakoid FtsH protease in Chlamydomonas. Plant Physiology. 191: 1002-1016